


正本溯源 ──生物親緣演化樹的脈理和論戰
王博群、吳宗賢、趙淑妙
由「樹」思考生物的親緣演化關係
早在18世紀中葉,生物學之父達爾文就首先運用演化樹(或生命樹)的觀念來描述物種的起源及多樣性(圖一)。想像一棵樹,它的地上部由樹幹及樹冠所組成, 樹冠內的枝條皆由樹幹發育 / 生出(也就是「起源」), 這表示樹冠的每一枝條(代表「物種」)起源於同一祖先,也就是有相同的「根」。而樹冠的枝條代表各類譜系家族,枝條上的側枝愈繁茂,則譜系內的物種多樣性愈豐富;反之,若枝條枯萎或折斷,則說明此類物種已滅絕, 若枝條冒出新的枝芽,代表演化出新的物種。親緣演化學的英文單字為「Phylogenetics」,是一個組合字,概念來 自「phyle」+「genesis」,前者為「譜系」或「家族」的意思,後者為「起源」或「發生」,所以這個英文單字很明確地指出親緣演化學的研究方向:物種譜系的起源與關係。
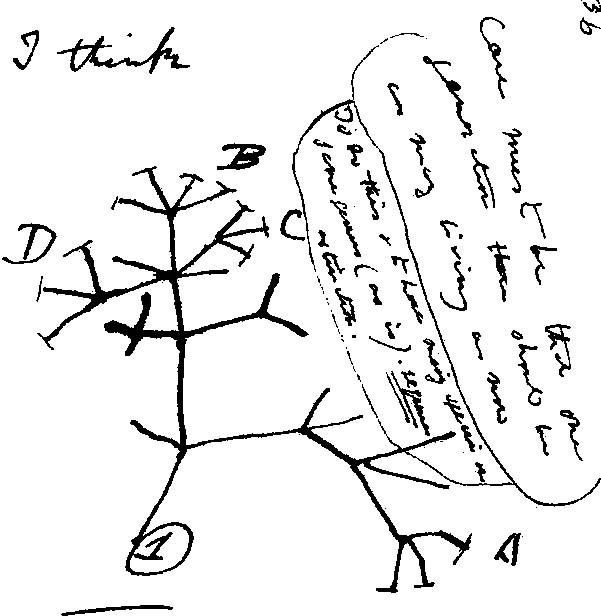
圖一、親緣演化樹的雛形,引用於達爾文在1837年手繪的手稿。
隨著生物學快速地進發展,當物種的DNA證據被應用在親緣演化樹的推論後,生物學家發現有「根」的演化樹已不能解釋複雜的生物演化史。如前所述,親緣演化樹是假設所有物種皆起源於同一祖先,亦即物種的遺傳物質皆由父母遺傳而來,即所謂的「垂直」遺傳,然而「水平」輸入遺傳物質的案例,在自然界中卻隨處可見。例如細胞藉由吞噬作用獲得被吞噬方的遺傳物質,最著名的案例是幾乎所有生物細胞中的粒線體及植物細胞中的葉綠體,它們分別是在生命演化的早期(約20億年前),由真核生物的祖先經由吞噬作用攝取變形菌及藍綠菌的遺傳物質而來。其實物種間遺傳物質的水平轉移比我們的想像還容易,且多發生於低等生物,如細菌及藻類等。有趣的是,水平轉移遺傳物質與物種的親緣從屬通常無直接關連。即使親緣關係非常疏遠的雙方,亦可互相轉移遺傳物質,例如無油樟(Amborella trichopoda)的粒線體擁有許多外源的DNA片段,其來源可能是與其有交互作用的藻類及苔蘚。有鑒於此,演化網或生命網(Phylogenetic reticulation)的進階概念應運而生,彌補了樹狀結構所無 法描述的雜交關係、基因水平轉移、遺傳重組等現象。演化網沒有末梢枝條,也就是說沒有任何一個枝條在分開以後,便永遠獨立於其他枝條。事實上,基因系統樹與物種親緣關係樹也常常發生不符合的狀況,但此類進階觀念非本文的重點,在此不予深究。
親緣演化樹的架構與型態
一、親緣關係
我們都有看過族譜(圖二),沒看過的讀者請查看家族神主牌背後是否有相關紀錄。在族譜裡,同一世代的人們被放在相同水平上,在同一世代裡,親兄弟姊妹與自己血緣關係最近,都是由同一對父母(祖先)所生;而堂表兄弟姊妹沒有共同的父母,父母和他們的兄弟姊妹則有相同的父母,也就是祖父母。從族譜可以總結出幾個重點:
1. 親緣關係是一種回溯的觀念。同一世代的個體其血緣關係一定可以追溯至共同祖先,同世代的個體間彼此不可能是對方的祖先。
2.親緣關係是一種集群(Cluster)的觀念。自己的親兄弟姊妹由父母所生,而堂兄弟姊妹則為叔叔嬸嬸所生,把自己及親兄弟姊妹視為一個集群,堂兄弟姊妹則為另外的集群,集群內的個體親緣關係緊密,而集群間個體的關係則較疏遠。我們把一對父母與他們的子女看成一個大一點的集群,另一對父母與他們的子女當成另一個大一點的集群,這兩個大一點的集群都擁有共同的父母(祖父母),如果將這兩個大一點的集群與祖父母一併來看,則又組成了一個更大的集群。想像一下用手抓住樹枝的一處,那麼從抓住的這一個點到從這個點擴散開的所有枝條(可以想像成這個點的子孫們)可被視為一個整體,這個整體在演化樹的結構上被稱作「分類群(Clade)」,意指可溯源至同一祖先的整體。
3.親緣關係是一種相對的觀念。想像一下你跟你的親兄弟姊妹,哪一位和你的堂兄弟姊妹的親緣關係較接近?答案是「一樣相近」。我們可以從圖二看出端倪,不論是你自己或是你的兄弟姊妹,與堂兄弟姊妹的關係都要追溯兩代才有彼此的共同祖先(祖父母);那麼你跟你的兄弟姊妹的關係較近或是跟你的堂兄弟姊妹較近?答案是跟兄弟姊妹較近,因為你跟兄弟姊妹只要追溯一代就可發現共同祖先(父母),而你跟堂兄弟姊妹的共同祖先則需追溯至二代(祖父母)。
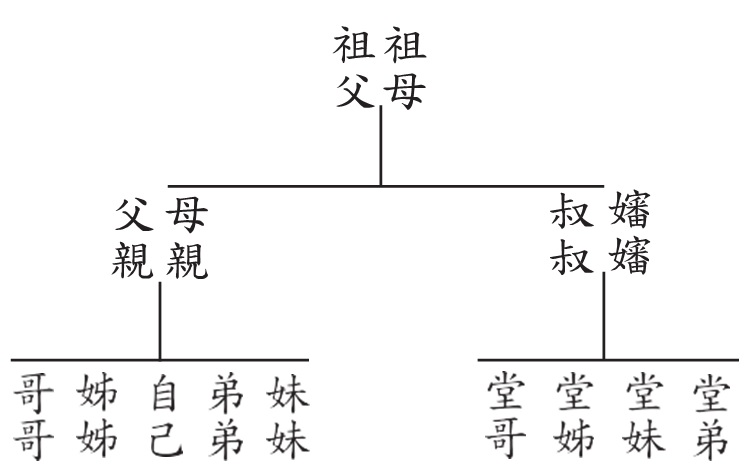
圖二、族譜示意圖,同一世代的人們會被放在相同的水平位置。
二、分支圖(Cladogram)
看完族譜後我們再談談分支圖。親緣演化樹常以分支圖表示,嚴格來講,分支圖不能代表親緣演化樹,親緣演化樹需有上跟下的從屬關係(祖先跟子孫)並提供時間軸予以對照。總之,我們可以把親緣演化樹簡化成分支圖的方式呈現,但分支圖不一定就代表親緣演化樹。分支圖將具有相同性質的物件集合在一起,描述它們的相對關係, 而所謂的同質性的物件不一定是遺傳物質,例如若是以財富區分人群所構成的分支圖,集群內的人擁有相似的資產,但他們卻沒有任何血緣關係。
常見的分支圖有幾種型態(圖三),雖然它們的形狀不同,但都表示類似的概念。以圖三為例,我們可以分成三個集群,分別為(A)、((B, C), D)、(E, F),將這三個集群的關係整合可得:(A,(D,(C,B)),(E,F)),也就是集群(E, F)與集群((B, C), D)的關係較近,但與A的關係較疏遠。我們只能從分支圖了解各物件的相對關係,所以說它是簡化的親緣演化樹表示方法。另外,分支圖中的節點(Node)是可以隨意180度旋 轉,旋轉不會影響物件的互相關係,以圖三(1)為例,物件在水平位置上由左至右為F、E、D、C、B、A,而圖三(2) 為B、C、D、E、F、A,但只要把節點α做180度旋轉,則物件的排列順序就變成一樣了。
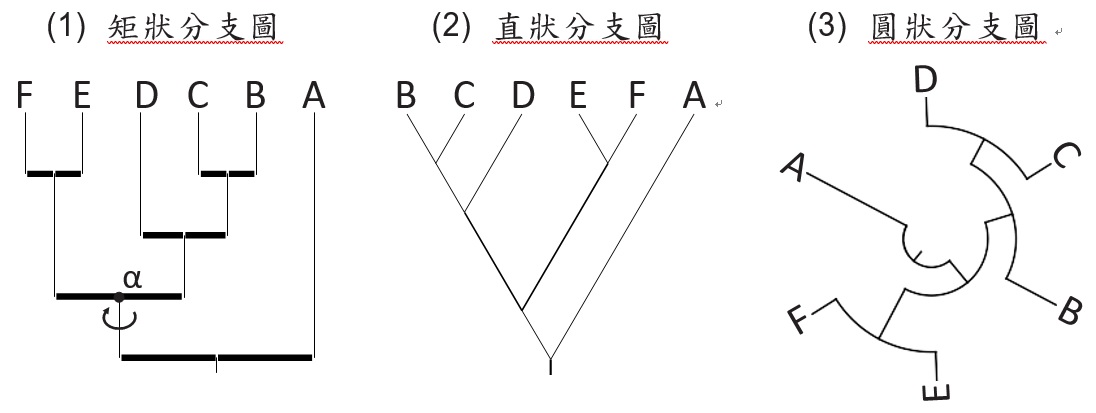
圖三、各種不同形狀的分支圖,但它們代表的物件相互關係都一樣。
親緣演化樹的組成元件
如上所述,分支圖只是簡化的親緣演化樹,因為分支圖內缺少一些重要的演化資訊。親緣演化樹則是由各種元件組成(圖四),我們將所要研究的物種或類階稱為「分類操作單元(OTU,是Operational Taxonomic Unit的縮寫)」。OTU可再分為內群(Ingroup)及外群(Outgroup),內群是我們要著手研究的物種,而外群是參考物種,有「定錨」的作用。親緣演化樹的線稱為枝條(Branch),圖四(1)內我們只關切垂直的枝條,水平枝條沒有意義。連接OTU的枝條稱為末端枝條,其餘則稱為中間枝條,枝條與枝條連接的點稱為節點,節點代表衍生出OTU的共同祖先。
親緣演化樹是有時間軸的,一般而言,所有OTU都是現生物種(Extant species),所以代表現在,因此由OTU回溯則代表過去的演化時間。OTU亦可能是滅絕物種(Extinct species),但常以虛線連結,且水平位置上會低於現生物種,如圖四(2)的F。若OTU的互相關係沒有足夠的資訊量支持,我們可針對親緣演化樹進行模糊解釋,如圖四(2)的B、C及D,雖然它們被歸類為親緣關係相近的一群,但它們之間的更詳細關係仍未被釐清。
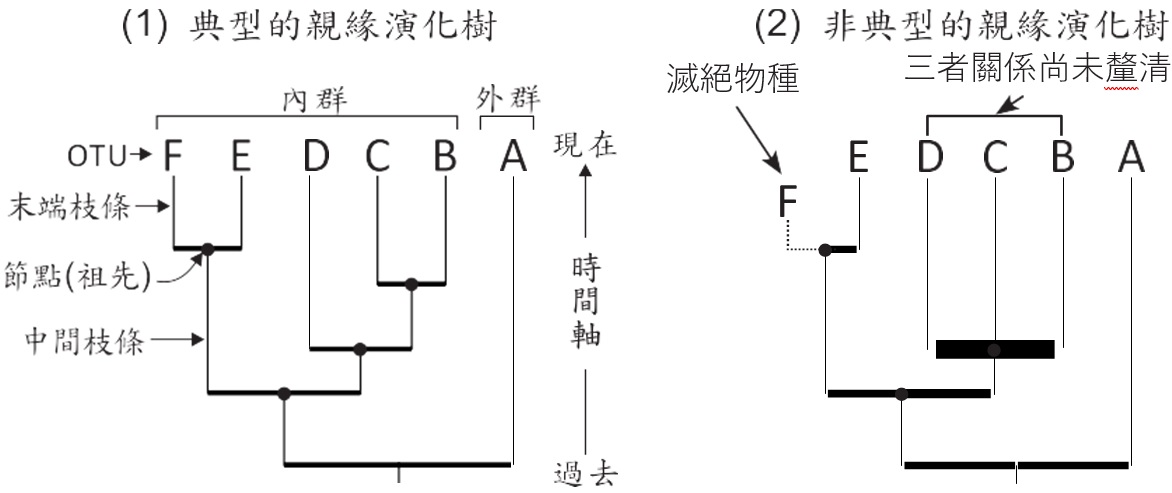
圖四、組成親緣演化樹的各項元件。
在有時間軸的情況下,枝條的長短代表物種分化後各自的演化時間。過去的研究已經知道各種生物的演化速率是不一樣的,單細胞生物通常有高於多細胞生物的突變率;老鼠的生命週期比人類短,因此老鼠的突變率亦高於人類。在OTU都是現生物種的情況下,我們可以用枝條的長短解釋DNA的核酸取代率,突顯物種具有不同的演化速率(圖五)。同樣地,若演化樹是以物種間彼此型態的相似程度來衡量,末端枝條的長度代表OUT被量化的特有型態特徵,中間枝條則表示OTU彼此間皆擁有的型態特徵, 所以中間枝條愈長,OUT彼此擁有愈多的相似特徵,而末端枝條愈長的OTU愈奇特,以圖五而言,OTU5是最奇特的物種。
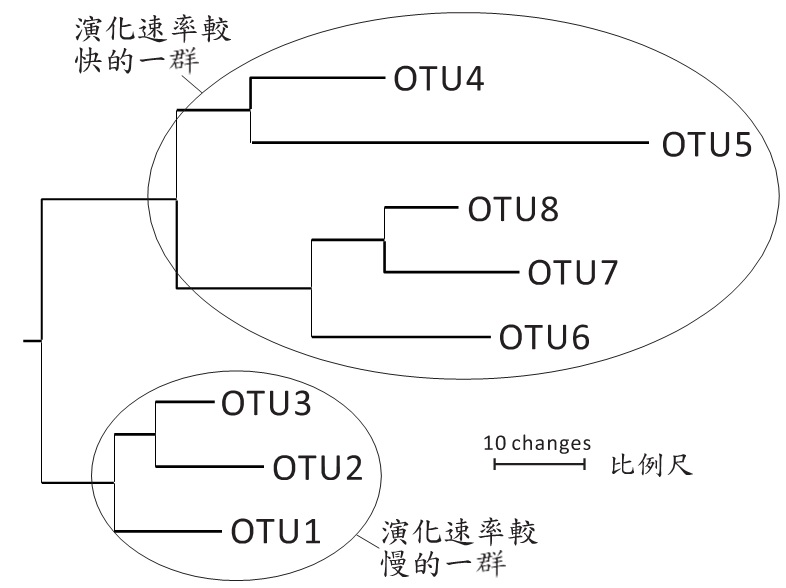
圖五、沒有時間軸的演化樹,枝條長度代表改變的程度。
判讀親緣演化樹常發生的誤解
現在我們對親緣演化樹有了初步的認識,但還是要小心翼翼地判讀,許多人往往會掉進一些錯誤的陷阱,以下便是幾個常見的誤解。
一、是否有主枝及旁枝
人們常常有這樣的誤解:演化的過程有一條主枝,而這條主枝的最末端是人類,就這個想法來說,所有的其他物種皆是從這條主枝衍生出來的,也因此人們覺得自己是地球上最高等的生物,特別是圖六(1)的直狀演化樹最易讓人產生這樣的誤解。然而如前所述,演化樹的節點是可以180度旋轉的,當我們握住節點並旋轉180度,結果如圖六(2)所示,我們便不會下意識覺得人類是最高等了。事實上,演化樹沒有主枝與旁枝的分別,生物亦沒有高等與低等的區別。回到本文篇首所謂的「樹」的概念,所有的枝條皆起源於主幹,這表示物種起源於同一祖先,然而樹冠的枝條代表各類譜系家族,而人類也只是眾多枝條的其中一支而已。
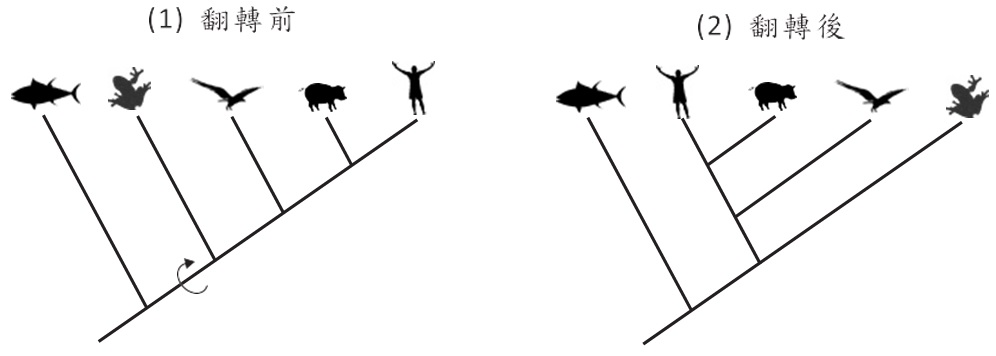
圖六、親緣演化樹沒有主枝與旁枝的區別,所以生物亦沒有高等與低等的區別。
二、排列愈靠近的物種是否親緣關係愈接近
再以圖六(1)為例,豬被排在人類的旁邊,有人因此會認為人類跟豬的親緣關係較近而與老鷹的關係較疏遠,這似乎合理,人跟豬都是哺乳類。同樣的邏輯,我們也可以說:青蛙跟魚類的親緣關係較近而與人類較疏遠,因為圖六(1)將魚類排在青蛙旁邊。但這個結論很明顯是錯的,魚類是以鰓呼吸,但人跟青蛙以肺呼吸,所以青蛙應該跟人類關係較近才對。那問題來了,為什麼用同樣的邏輯卻產生不同的結果?這是因為我們用了錯誤的邏輯。我們解讀演化樹要順著枝條回溯,從人類的枝條回溯,第一個遇見的是人與豬的共同祖先,再來才是人與老鷹的共同祖先; 同樣地,若是從青蛙的枝條回溯,先碰到青蛙跟人的共同祖先,然後才是青蛙與魚類的共同祖先,所以正確的描述是:青蛙與人類關係較近而與魚類則較疏遠。更何況如前所述,親緣演化樹的各個節點可以180度旋轉,旋轉後的圖六(2),其物種水平位置排列跟圖六(1)完全不同,難道我們又要改變看法?當然不是。所以判讀演化樹一定要隨著枝條回溯,才不會發生錯誤的判讀。
三、是否有最原始的物種
當我們看到某一枝條都沒有分歧且被放在親緣演化樹的最外圍時(圖七(1)),往往會誤判A物種是最原始的, 因為以時間軸來看,A物種出現的最早,存活的年代最久,但這種判斷會陷入取樣不足及沒有考慮物種滅絕的陷阱。例如,建構親緣演化樹的當下,並未取樣與A物種親 緣關係近的生物,使A物種特異於其他物種,造成A物種已存活很久的假象。或許有人會質疑:如果是在物種取樣上非常完備的情況下,是否能說A物種是最原始的呢?答案 仍舊不行,因為要考慮物種滅絕的可能性。我們已知地球發生過幾次的物種大滅絕,每次都有超過90%以上的生物絕滅,當已滅絕的物種被併入親緣演化樹時(圖七(2)), 我們就不會覺得A物種是最原始,畢竟從時間軸來看, B、C、D、E物種的出現及存活時間皆比A物種久遠。事實上,親緣演化樹不能提供絕對的信息,而「最古老」或「最先進」就是絕對的信息。親緣演化樹僅告訴我們誰跟誰的親緣關係較近或較遠,在圖七(1)中,我們可以說D物種的親緣關係與E物種較近而與A物種較疏遠,因為順著D 物種往回追溯會先碰到D與E物種的共同祖先,再繼續追溯才會碰到D與A物種的共同祖先,以時間軸來看D與A物種共同祖先出現的年代確實早於D跟E物種的共同祖先。
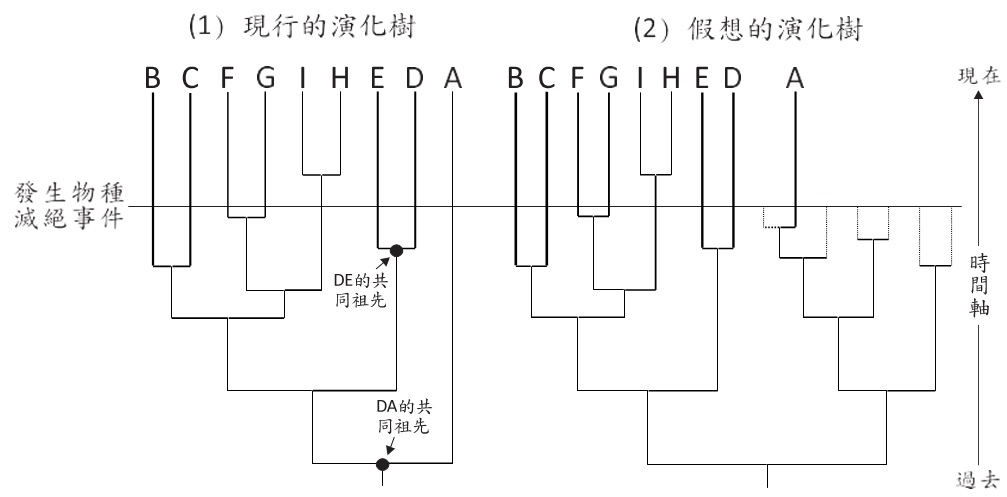
圖七、親緣演化樹沒有最原始的物種。當考慮物種滅絕事件後,我們發現所謂的最原始物種,是對物種親緣關係的錯誤解讀。
四、改變僅發生在節點嗎
親緣演化樹的節點是我們特別感興趣的信息,那麼節點代表甚麼呢?代表了物種在這個時間產生分化嗎?如果是這樣,也就是說在節點所在的時間點,生物的性狀忽然發生了很大的歧異嗎?必須要澄清的是:生物是群體,我們假設兩個物種的共同祖先有兩個群體,這兩個群體平時都有進行基因交流,但基因交流在演化的某個時間點中斷了,最常見的原因是地理的隔離。中斷基因交流使兩群體獨立演化,而隨著遺傳因子突變的不斷累積,最終造成生殖隔離,演化出兩個新的物種。簡而言之,節點就是共同祖先群體內發生生殖隔離的時間點。如前所述,改變(或是突變)是不斷地累積的,而節點可視為物種分化的爆發點。有趣的是,人類對動植物的選拔及育種也會促進物種分化,在不斷地追求「純」種品系的要求下,某些品系與野生種將可能產生生殖隔離,進而加速物種演化。
親緣演化樹是假說
親緣演化學是以現有的證據(DNA或形態學證據)推論過去發生的事件(親緣關係事件),追溯的事件小至親代的關係,例如人們的DNA驗親;大至整個生物界的親緣關係,例如近期很流行的地球生命樹(Tree of Life)。但在此要特別強調:我們無法回到過去,目睹生物的演化過程,我們只能像偵探一樣,用手上現有的資料來推論,而且事件發生的越久遠,推論的困難度越大,因為在時間長河的淘洗下,證據清晰度往往被消磨大半。也就是說推論近代親緣,如父子間的驗親,其可信度很高,但推論跨大類階的物種關係,如生物界的科與科之間的關係,或目與目之間的關係,使用不同DNA證據,或不同的演算法,推論的結果可能會不一致。
以裸子植物的親緣演化關係為例,諸多親緣關係假說已被提出,學界對裸子植物親緣關係的爭論,紛擾近30年卻仍未定論。現生的裸子植物可分成五個類群,分別為蘇鐵、銀杏、柏類、松類、買麻藤類。若加上被子植物,則組成所謂的種子植物(圖八(1))。蘇鐵及松柏類我們在校園或植物園常見,銀杏相信大家也不陌生;但買麻藤在臺灣卻很少見,這類植物的原生地在東南亞及其他熱帶國家,其種子常在當地被視為食物。如圖八(1)的照片所示, 買麻藤看起來如同被子植物,葉子擁有葉脈構造,具有如同藤蔓般的纏繞莖可攀爬其他植物,但買麻藤具有毬果, 種子無種皮,這些特徵又與其他裸子植物相似。早期的生物學者是以形態學為證據,得到買麻藤與被子植物親緣關係較近而與其他裸子植物較疏遠的假說,也就是所謂的「生花植物假說(Anthophyte hypothesis)」,但這個假說卻被以DNA為證據的分子演化生物學研究所推翻。
然而,以DNA 為證據的演化學研究也衍生出各種不同的假說,如圖八(2) 所示,「買麻藤松柏類假說(Gnetifers hypothesis)」,這一派假說支持買麻藤與松
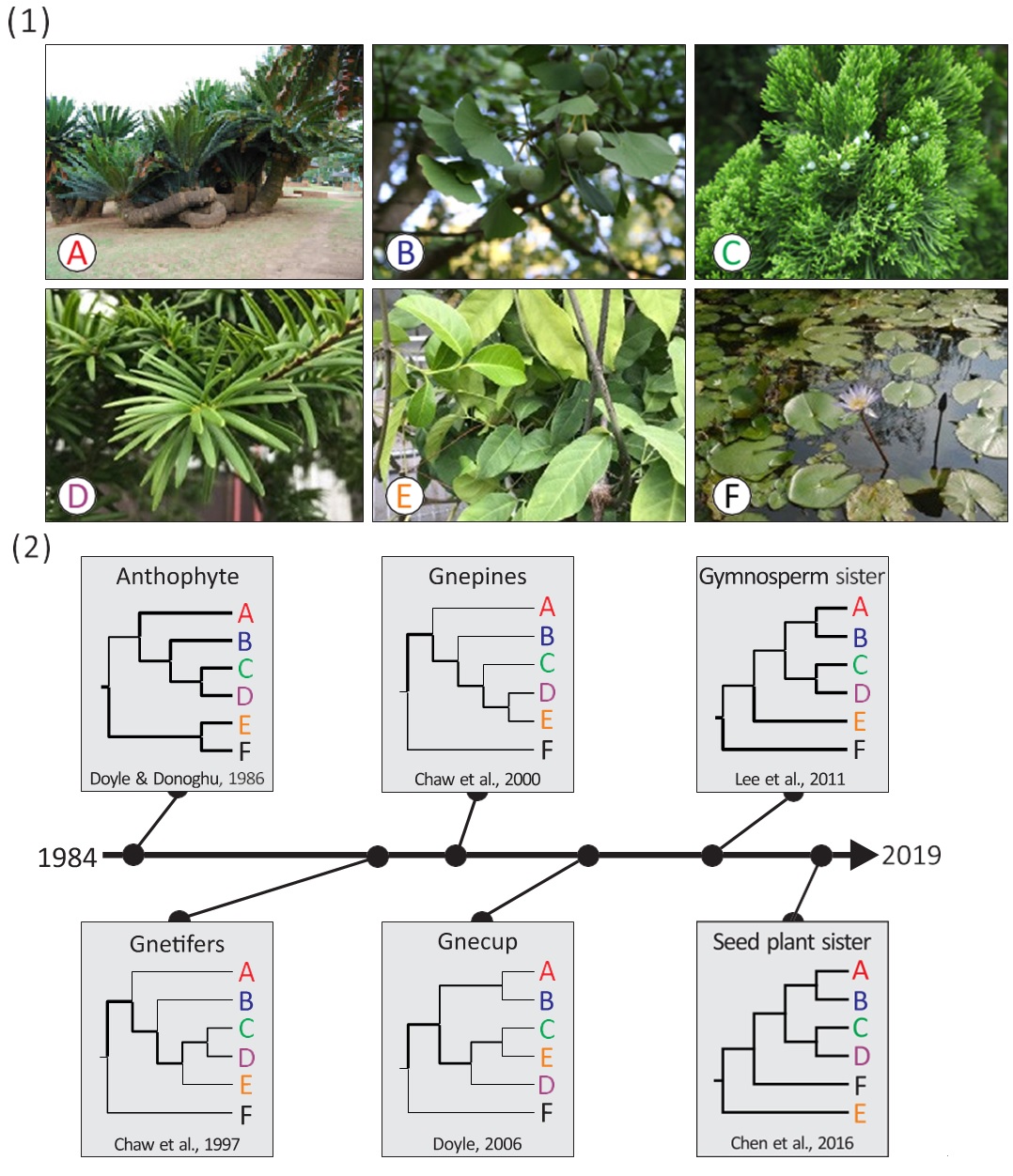
圖八、裸子植物的親緣演化假說。(1)現生的六大種子植物類群:(A)蘇鐵;(B) 銀杏;(C)柏類;(D)松類;(E)買麻藤類;(F)被子植物。(2)過去被提出過的裸子植物親緣演化假說。
柏類是關係最近的姊妹群;另一派假說則認為松類與買麻藤關係較近而與柏類的關係較疏遠,即所謂的「買麻藤松類假說(Gnepines hypothesis)」;「買麻藤柏類假說(Gnecup hypothesis)」則抱持相反觀點,支持柏類與買麻藤是關係最近的姊妹群;「裸子植物姊妹群假說(Gymnosperm sister hypothesis)」則認為買麻藤是其他裸子植物的姊妹群;「種子植物姊妹群假說(Seed plant sister hypothesis)」則推論買麻藤是其他種子植物的姊妹群。在此需特別聲明,較晚發表的假說不代表其可信度較高,所有的假說都是「推論」,也是可以被挑戰的。哪天有位生物學家發現新的證據,或是對舊證據提出新的詮釋,新的親緣演化假說就會被提出。所以說親緣演化樹所推論的物種親緣關係僅代表各項假說,只要生物學家提出合理的證據及詮釋,能說服其他學者,假說就可以被接受。
結論
常聽人說人類是從猴子演化而來的,但這種直線式的思考是不正確的。較精確的說法是:猴子跟我們是兄弟姊妹的關係,我們跟猴子擁有共同的祖先,因此我們沒有比猴子高等。「樹」狀的思考催生出親緣演化樹的應用,也更能準確地描述物種的親緣關係。讀者若對親緣演化學感興趣,想更深入地了解其學說及研究方法,不妨研讀Page & Holmes博士的著作Molecular Evolution: A Phylogenetic Approach及Dan Graur博士與李文雄院士撰寫的Fundamentals of Molecular Evolution。
後 記
我們正處於DNA定序技術蓬勃發展的時代,藉由資源共享, 得以獲致更豐富而完善的分子資料。許多傳統理論 / 見解,正等著我們以大數據、生物資訊、新演算法去重新檢驗,歡迎讀者與我們一同加入探索物種起源的行列。
作者簡介

王博群
臺灣大學地理與環境資源學學士,現就讀中央研究院生物多樣性國際研究生學程(TIGP Biodiversity Program),興趣是分子演化。參與此本科普書的撰寫是他的首次經歷。

吳宗賢
臺灣大學森林學博士,中央研究院生物多樣性研究中心博士後研究學者。農家子弟,平常熱愛山林探索。

趙淑妙
中央研究院生物多樣性研究中心特聘研究員兼主任。美國杜蘭大學生態與演化學博士。專長植物親緣演化學、胞器基因體學、及核基因體學。研究物種以裸子植物及被子植物為主。閒暇時熱心公益活動,為婦女團體發聲,鼓勵女性從事科學研究。翻譯《自私的基因》一書,深獲好評。
