



變調的生命之歌── 臺灣溪哥被盜用「身份證」的一頁滄桑?
王子元、黃世彬

圖一、採自淡水河的日本溪哥(平頜鱲)。(王子元 攝)
1980年代是水產養殖興盛的年代,也是臺灣經濟命脈起飛的年代。經濟與生態間的對立,似乎也從此刻開始。
家家戶戶喜愛的香魚(Plecoglossus altivelis)和溪哥(原為鱲屬Zacco,現生臺灣原生溪哥被歸類為馬口鱲屬Opsariichthys),曾經普遍分布在臺灣北部的河川裏,是 釣客們的最愛之一,更是山產店常見的下酒菜餚。酥酥脆脆的油炸溪魚,也是居民補充鈣質、經濟實惠的營養品。然而,由於工業汙染環境日趨嚴重,造成河川水資源的惡化,加上電魚、毒魚、炸魚等粗暴的捕獵方式,使得河川有水、水裡有魚的景色逐漸自上一代的記憶中消逝。
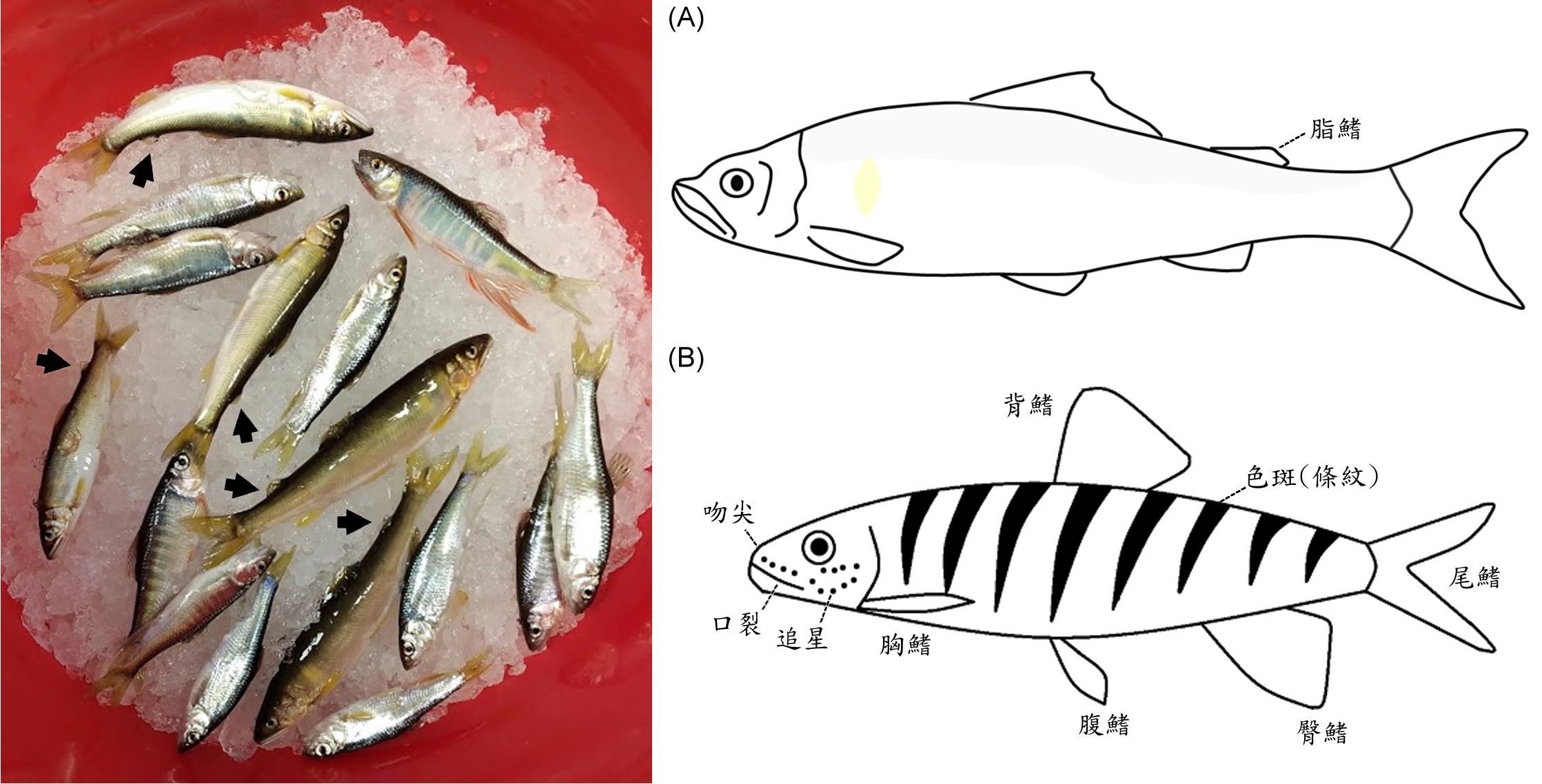
圖二、新店溪的香魚和溪哥。香魚體色偏黃,背鰭和尾鰭之間有個柔軟的肉質鰭(脂鰭,左圖箭頭處)(A)。溪哥體色銀白,成熟雄魚具有數條橫斑和追星(B)。(王子元 攝)
臺灣當年的水產養殖技術是國家的重點研究工作,河川復育的先驅觀念也是從水產試驗研究單位開展。放流魚苗是當年常見的復育方式,也是縣、市政府積極投入的策略。例如,政府機構意識到臺灣香魚族群的滅絕,在1960 年代初,從日本引進同物種的香魚受精卵,試圖重現臺灣的香魚族群。終於在1977年蕭世民等人透過水產養殖技術,成功地孵化香魚苗,自此陸續將為數眾多的香魚苗放流到新店溪上游、石門水庫、萬大水庫、德基水庫等地, 藉以維持有香魚族群存在的風貌。

圖三、臺灣河川中的日本香魚。(周銘泰 攝)
隨著生物技術的進展,以及生態系平衡的新觀念產生,生物多樣性的研究重新定義了物種間的親緣關係和族群間的遺傳差異。國人開始意識到即便是外表長相相同的物種,在地理隔離的自然歷史中,族群間也存在著具有指標意義的遺傳變異,甚至有可能存在著未知的新物種。昔日的放流方式,可能造成族群的獨特性逐漸消失。就像是移民和原住民的婚姻關係,混合的血統改變了昔日不通婚的獨特外型。臺灣的鳥類也曾發生這樣的案例──「雜頭翁」是東部特有種的烏頭翁(Pycnonotus taivanus)和分布西部的白頭翁(Pycnonotus sinensis)的後代,其羽色及體型兼具兩個物種的特徵和特性,並具備生育能力;這兩種被中央山脈隔離了十幾萬年的生物,卻因公路建設、人為放生與自然棲息地的開發等非天然因素,打破了地理阻隔的藩籬,兩個物種交配產生了不同形態的雜交後代。隨後,強勢入侵東部的白頭翁已經壓縮了烏頭翁的生存空間。

圖四、臺灣西部的白頭翁和東部的烏頭翁雜交,因而產生不同表型的雜頭翁子代。(雜頭翁C:官建維 攝;其他各圖:詹仕凡 攝)
當人們吃著山產店的油炸溪魚時,是否曾想過這「山產」來自何處呢?
臺灣的溪哥,早期被學者認為共有三種,即與日本同名的平頜鱲(Zacco platypus)與丹氏鱲(Z. temminckii) 以及臺灣特有種粗首鱲(Z. pachycephalus)。1986年, 曾晴賢指出丹氏鱲是錯誤鑑定,並不存在於臺灣。而臺灣的平頜鱲被部分學者認為應該是長鰭鱲(Z. evolans) 的錯誤鑑定。直到2005年,陳義雄與張詠青合著的《臺灣淡水魚類圖鑑》始將臺灣產的原生溪哥歸到馬口鱲屬, 並且在2009年發表一個產於臺灣南部的新種──高屏馬口鱲(Opsariichthys kaopingensis)。至此,臺灣所產原生溪哥物種的分類已經釐清,除了高屏馬口鱲,臺灣原生的溪哥尚有分布於西部與東北部的粗首馬口鱲(O. pachycephalus)以及北部的長鰭馬口鱲(O. evolans)。
2006年,馬國欽等人利用分子親緣鑑定技術,意外發現臺灣的淡水河族群存有和日本琵琶湖同源的平頜鱲(本文之後稱日本溪哥)族群,恰巧和引進臺灣的日本香魚來源相同。然而,當時文獻上並無明確的放流記載;因此, 有專家推測日本溪哥的魚卵極可能夾雜在日本香魚卵當中,意外地被放流到臺灣的新店溪一帶。也有可能是當時流行飼養溪魚的水族養殖業者引進臺灣,或是後來民間人士的宗教放生活動所致。亦有學者認為臺灣的確尚存著本土的平頜鱲族群。就像香魚和雜頭翁的雜交例子,這也是一個需要釐清的歷史事件。
臺灣產的溪哥無論外形與遺傳上均與日本溪哥有所不同。2010年王晨帆、王弘毅等學者根據型態以及DNA的研究發現:臺灣的河川存在日本溪哥與臺灣兩種溪哥的雜交個體。人為引進的日本溪哥, 竟能夠和臺灣原本互不雜交的兩種溪哥,分別產生出雜交後代!進一步翻閱日本水產文獻, 在日本各地已陸續發現有平頜鱲與其他近緣屬(種)的雜交個體。這些非自然現象,主要起因於人為水利工程,改變了適合生存的棲地溫度或發生棲地匯合,增加了物種間雜交繁殖的機會。
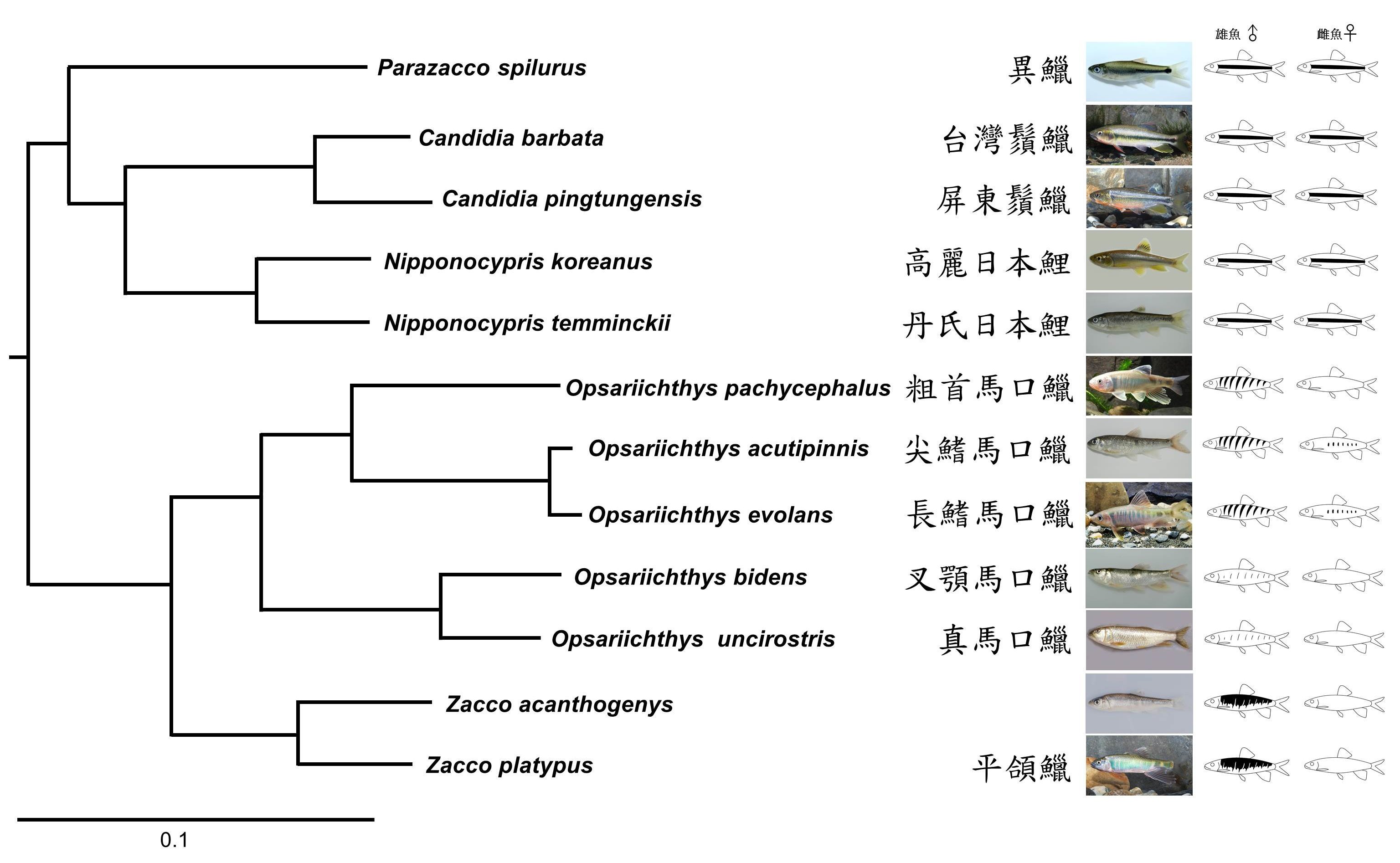
圖五、利用全粒線體DNA序列重建溪哥和馬口魚類的親緣關係樹(左)和體側婚姻色的斑紋型態(右)。(黃世彬 製)
我們推測造成這樣的雜交個體,也許和溪哥特殊的生殖行為息息相關。許多魚類在繁殖的季節,成熟的雌雄個體會產生配對的行為, 配對成功的雄魚會巡弋在雌魚的周邊,驅趕其他的雄魚, 以確保繁殖過程中,能夠產生自己的後代。溪哥就是這類典型的例子。成熟的雄魚具有數條不同顏色的相間條紋, 被稱為「婚姻色」,是雌魚用來辨識雄魚是否健康成熟的特徵。配對成功的雄魚,會尾隨在尋找產卵地點的雌魚,並不時追逐驅趕靠近雌魚的其他雄魚。這些「未配對成功的雄魚」被稱為配對行為中的「衛星個體」。這些衛星個體會藉機靠近正準備交配的雌魚,趁著配對的雄魚和雌魚共同搧尾交配,無暇他顧的瞬間,靠近雌魚將牠們的精子也同時撒下。由於魚類是體外受精,此種趁亂而入的投機行為,是可製造受孕的機會,將自己的遺傳基因繁衍下去。

圖六、疑似平頜鱲和粗首馬口鱲的雜交個體的雄魚(上)和雌魚(下)。(周銘泰 攝)

圖七、具有婚姻色的平頜鱲成熟雄魚(上)。(王子元 攝)

圖八、骰子相機拍攝到長鰭馬口鱲(照片左側)和平頜鱲(照片右側)在同一棲地產卵。(王子元 攝)
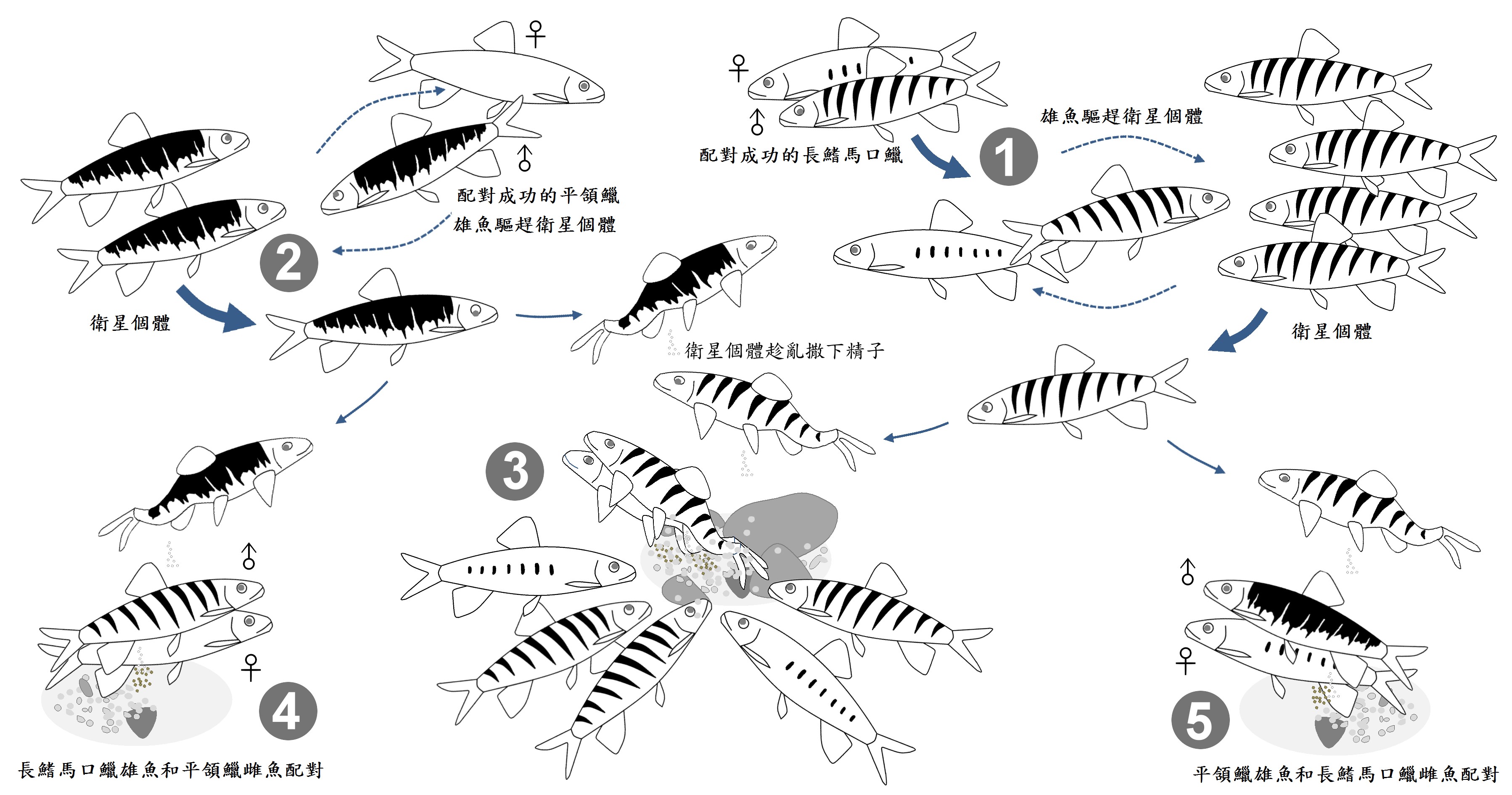
圖九、長鰭馬口鱲和平頜鱲的雜交事件簿。(王子元 繪製)(1)配對成功的長鰭馬口鱲,雄魚會驅趕其他靠近的雄魚;(2)配對成功的平頜鱲雄魚也會驅趕其他靠近的雄魚;(3)產卵中的長鰭馬口鱲上方會出現衛星個體趁亂撒下精子,後方則會有其他個體尾隨;(4)會有少數配對發生在長鰭馬口鱲的雄魚和和平頜鱲的雌魚;(5)將近有三分之一配對發生在平頜鱲的雄魚和和長鰭馬口鱲的雌魚。上述多種組合配對和衛星個體參與的狀況,產生了許多雜交的機會。
圖十、樣區採樣記錄。(王子元 攝)
雜交個體是否因為這樣的特殊行為而產生的呢?2018 年為了證實這個現象,我自費買了一個可以水下攝影的骰子相機,並帶著小型攝影機到了王晨帆曾發現雜交個體的「事故現場」,試圖拍攝這樣的雜交行為。骰子相機是一款用於運動登山記錄的小型攝影機,也有人會買來黏在安全帽上做為騎車記錄器使用;似乎不曾有人拿來記錄水下的生態,沒想到用在這次觀察實驗中,非常實惠好用!就在第三次前往現場記錄時,我從手持攝影機發現一對正在配對產卵的平頜鱲上方,出現了長鰭馬口鱲的雄魚側身搧尾的標準動作!難掩興奮的當天,光手持攝影機就記錄到三次衛星個體表現出關鍵行為的畫面,而且不只發生在平頜鱲,平頜鱲搧尾的行為也發生在配對產卵的長鰭馬口鱲上方。這是野外首度證實兩種溪哥在同一棲地產卵,衛星個體可能是造成雜交後代的關鍵。下午回到實驗室,我們興奮地趕緊檢閱水下攝錄的骰子相機,畫面同樣出現溪哥配對和衛星個體參與異種交配的清晰畫面!經再三觀看拍攝到的幾段畫面後,證實了不同種交配事件的存在。
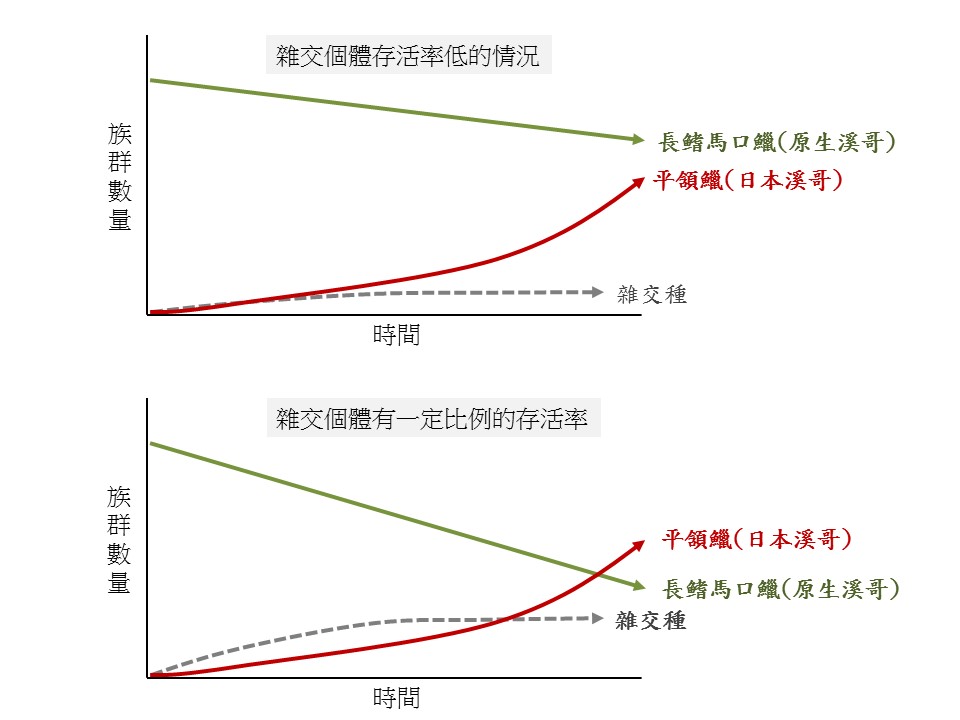
圖十一、外來種的雜交行為對原生種族群變動的影響。(王子元 製)
不同種間的交配行為,究竟是隨機發生的事故,或者是有選擇性的偏好呢?三個月後的暑假,四位參與中央研究院生物多樣性研究中心舉辦的大學生暑期培育計畫,來自高醫、東吳和臺大的學生,加入了這個觀察研究,讓我們得以進一步探討這個「事故」。在中心研究經費的挹注下,我們增購幾組骰子相機,以便拍攝更多的畫面,進一步統計分析。
經過多次現場觀察記錄和影像辨識的分析工作,平頜鱲和長鰭馬口鱲間的雜交行為,竟然比想像中的更加複雜頻繁!除了衛星個體造成的雜交事件外,在觀察到的73組配對中,將近有三分之一的配對是發生在平頜鱲的雄魚和和長鰭馬口鱲的雌魚。若再加上衛星個體的種間雜交行為,只剩下43%的產卵事件是屬於原生種的長鰭馬口鱲。假設雜交個體的存活率較低,則種間交配頻繁的繁殖行為,可能會降低原生種的成功繁殖率,因而導致原生種後代數量的顯著減少。再者,若雜交個體有一定比例的存活率,如同雜頭翁可以繼續繁殖產生後代,可能會使原生種的族群數量相對的減少。人為引入的日本溪哥,或許也會如同強勢入侵東部的白頭翁一樣,取代東部特有的烏頭翁,成為主要族群。因此,未來有必要進一步瞭解平頜鱲與馬口鱲魚類的雜交行為與成因,以及監控外來種的平頜鱲族群數量及分布範圍。
「有心栽花花不開,無心插柳柳成蔭。」似乎精確地反映了外來種入侵的典型事故,這成語也同樣反應在臺灣的河川生態現況。例如,過去封溪護魚的立意雖然良好, 遇上了近年來的野溪整治工程,甚至人造親水公園,最終導致「保育吳郭魚」有成,而原生溪魚的族群卻因棲地生態被破壞而銳減,甚至走上滅絕的結果。才會因而產生需要透過人為放流魚苗的手段來達到物種復育的目的。日本溪哥與臺灣溪哥產生雜交個體,正是放流復育所產生的後遺症。事實上,維持棲地多樣性的保育工作,是最省錢也是最能達到保育目標的方式。政府若能做好淡水河和愛河的污染排放管制等淨水工程,必能落實在縣市的「河川整治」工程上— 在兼顧防災需求下,營造出臺灣溪魚能夠生存的特有溪流環境。如同臺灣創作女歌手潘美辰在《我想有個家》的歌詞﹕「我想要有個家,一個不需要華麗的地方,在我疲倦的時候,我會想到它。我想要有個家,一個不需要多大的地方,在我受驚嚇的時候,我才不會害怕。」何時能讓臺灣的溪魚也能有個家,不要只是羨慕吳郭魚,受傷後可以回家。別讓臺灣的溪魚只能孤伶伶地尋找牠已破碎的家園—那個過去記憶中的青山綠水!
[註] 本文「平頜鱲」指的是原產於日本,後引進臺灣的日本溪哥。
生態小學堂

三種溪哥的形態特徵。(黃世彬 攝製)
a)長鰭馬口鱲雄魚; b)長鰭馬口鱲雌魚; c)平頜鱲雄魚; d)平頜鱲雌魚; e)粗首馬口鱲雄魚; f)粗首馬口鱲雌魚。
共棲於臺灣北部淡水河流域的粗首馬口鱲( O . pachycephalus)、長鰭馬口鱲(O. evolans)、外來種的平頜鱲(Z. platypus)等3種雄魚當中,粗首馬口鱲、長鰭馬口鱲、平頜鱲的亞成魚體側色斑差異小,不易區別;成魚在繁殖季時的體側色斑則具有不同顏色及斑紋型態。以共棲於淡水河的平頜鱲(Z. platypus)和長鰭馬口鱲(O. evolans)為例,有四個顯著的形態特徵差異—不同的胸鰭長度、魚嘴邊突起的追星型態、雄魚體側色斑型態、雌魚吻尖顏色差異。然而,僅有兩個形態特徵可以區分這兩種雌魚(圖b、d)。首先,長鰭馬口鱲雌魚會有幾條黃綠色條紋(5),平頜鱲雌魚的條紋則為模糊的淡黃色或灰色(10)。其次,長鰭馬口鱲雌魚的吻尖端通常為灰綠色(6), 但平頜鱲雌魚通常為橘紅色(11)。
長鰭馬口鱲的背部底色為黃褐色,成熟雄魚的身體具有11∼13條垂直的綠藍色條紋(1),胸鰭相當長,最大長度可達到成熟雄性的腹鰭基部(2),但雌魚的條紋較短(5),胸鰭也明顯較短。在繁殖季節可以清楚地觀察到性別外觀的差異(圖a、b)。成熟雄魚在頰部與吻部上會分布顆粒狀追星(3)、尾柄呈現黃綠色(4);而雌魚則沒有這兩個特徵。此外,成熟雄魚的部分胸鰭、背鰭和臀鰭具有紅色斑,這些鰭膜在成熟雌魚通常為黃色。
平頜鱲在背部區域也具有黃褐色底。但是身體除了有幾條垂直的灰藍色條紋,部分條紋在成熟的雄魚中則融合成寬帶(7)。雌魚則呈現模糊的淡黃色或灰色條紋(10)。平頜鱲的胸鰭明顯較短,最大長度不及腹鰭基部(8)。在繁殖季節也可以清楚地觀察到性別外觀的差異(圖c、d)。成熟的雄魚吻尖是灰黑色(9),在臉頰和鼻子邊會有縱列追星;而雌魚則沒有追星,吻尖是橘紅色(11)。此外,成熟 雄魚的部分胸鰭、腹鰭、背鰭和臀鰭具有紅色鰭膜,這些鰭膜在成熟雌魚通常為黃色。
粗首馬口鱲有幾個關鍵特徵明顯與其他兩種不同(圖 e、f)。第一,粗首馬口鱲的口裂較大,後方可達眼球中點下方,上顎前端具有明顯的缺刻(12)。第二,粗首馬口 鱲的雌雄差異明顯—成熟雄魚在頰部與吻部上具有顆粒狀追星、體側的條紋為藍綠色(13);鰓蓋下方、腹鰭、胸 鰭、臀鰭等多處會有鮮紅色。雌魚則通體銀白,背部灰綠,體側具有不明顯的淺灰色橫斑(14),鰭膜呈淡黃色。
後 記
就像是疫苗開發的期程,物種保育是冗長艱辛的路程。相反地,隨意放生或丟棄外來種的危害,卻像肺炎流感的輕易咳嗽, 擴散得非常迅速。物種保育成功與否,有賴於全民高度地自主管理!要有「公德」,才會有「功德」。
作者簡介

王子元
中央研究院生物多樣性研究中心研究副技師,清華大學生命科學博士。專長為分子生態與演化、魚類基因體與奈米孔定序。主要研究微生物功能基因演化與應用、淡水魚類的親緣地理學、生殖行為與基因體研究。

黃世彬
中央研究院生物多樣性研究中心博士後研究學者,生物多樣性研究博物館的動物標本館資深經理。專長為魚類系統分類、魚類分子親緣以及魚類生態學。主要研究對象為東亞與東南亞的淡水與河口魚類,近來也推廣科學知識與博物館藏資源給社會大眾與學生。

保存在中研院生物多樣性研究博物館的三種溪哥早期採集標本:平頜鱲(上)(標本號ASIZP0074651)、粗首馬口鱲(中)(ASIZP0054186)、長鰭馬口鱲(下)(ASIZP0062093)。分別採集於2002年、1968年以及1997年。

