見微知著──由雛鳥新生絨羽一窺現代鳥類演化的奧秘
李文雄、陳志冠
鳥是分布很廣的陸生脊椎動物,擁有多樣的體型和豐富的顏色,至少已經有一萬種的鳥被記錄。鳥類與其他脊椎動物的不同之處,在於鳥類擁有羽毛、沒有牙齒、快速的新陳代謝以及輕巧但結實的骨骼。尤其是複雜多變的羽毛有利於鳥類佔據不同的生態棲位(ecological niches)。因此,鳥類很適合作為模式物種,來研究動物如何在多樣的環境中生存與適應。
羽毛在鳥類的演化過程中表現出高度的多樣性與複雜性。例如,由羽枝和羽小枝組成的柔軟絨羽可以保暖,而帶有羽軸的廓羽(輪廓羽毛)則可保護身體和吸引異性, 再者,不對稱的飛羽適合氣體動力學的原理,有利飛翔(圖一)。因此,羽毛的多樣性,很可能是鳥類發展出高度多樣性的最大原因。
不同鳥類在剛孵化的雛鳥(幼鳥)階段,成熟度呈現明顯的差異,這個現象被歸納成「早熟—晚熟頻譜(altricial-precocial spectrum)」(表一)。鸚形目(Psittaciformes)和雀形目(Passeriformes)等鳴禽在剛 孵化的時候,全身幾乎沒有羽毛而且眼睛仍未睜開,屬於晚熟鳥。相反地,諸如雞形目(Galliformes)和雁形目(Anseriformes)雛鳥的狀態則與成鳥相近,羽毛包覆全身而且眼睛張開,屬於早熟鳥。雛鳥的成熟度決定了牠們需要親鳥照料的程度,以及牠們適應環境的能力。
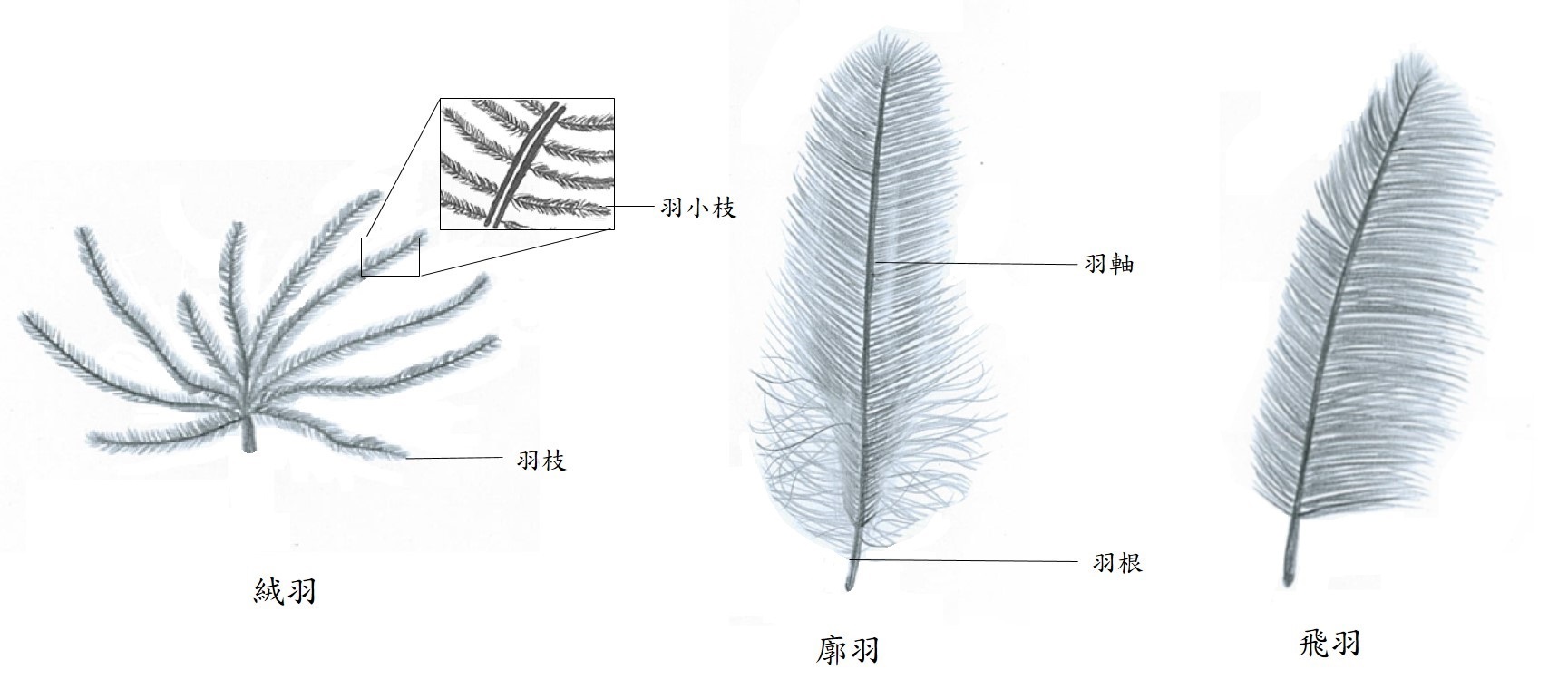
圖一、絨羽、廓羽及飛羽的構造。(戴嘉葳繪圖)
雛鳥新生絨羽的形態與生長部位會依據鳥種的發育特性(早熟鳥抑或晚熟鳥)有所不同,因此羽毛是用於辨識早熟鳥與晚熟鳥的主要特徵。這些發育特性和環境適應的模式,似乎可以幫助我們了解鳥類的演化歷程,然而相關研究至今甚為有限。本文中,我們首先以最近發表的鳥類親緣樹分析「早熟—晚熟頻譜」的演化模式。其次,探討雛鳥新生絨羽的演化歷程,藉此一窺現代鳥類演化的奧秘。
早熟鳥與晚熟鳥的定義及牠們的演化
早熟鳥與晚熟鳥雛鳥的差異,被認為和適應環境有關。然而如何定義早、晚熟鳥仍存在爭議。在棲息地的選擇上,大多數的晚熟鳥傾向於在樹上或高處築巢,牠們的雛鳥需要在巢上成長一段時間才能獨立,進而離開巢穴。相反的,大多數的早熟鳥傾向於在地面築巢,雛鳥孵化後不久就可以獨立並離開巢穴。雛鳥的一些形態和行為特徵,常被用於辨識早、晚熟鳥(表一),例如新生絨羽的多寡及種類、活動能力、親鳥的照護程度、獨立覓食的能力、待在巢中的時間、孵化時開眼或閉眼以及與親鳥之間的互動情形等(表一)。
我們將「早熟—晚熟頻譜」對應到最近發表的鳥類演化樹上(圖二)。由於可得的資料有限,我們僅使用四種雛鳥發育模式:早熟、半早熟、半晚熟、晚熟。過去的化石及系統發育學(一種研究生物個體或群體之間的演化歷史和關係的學科)的研究支持了鳥類從早熟鳥演化到晚熟鳥的趨勢,我們的結果也符合這個推論(圖二)。然而, 從早熟鳥到晚熟鳥的演化,在整個鳥類演化史中曾發生過數次,僅列載細節如下:古 顎 下 綱 ( P a l a e o g n a t h a e ) 和 雞 雁 小 綱 (Galloanserae)是兩個最古老的鳥類譜系,所有的成員都屬於早熟鳥(圖二)。新鳥類(Neoaves)則包含了大多數的現生鳥類譜系,主要有:夜鷹及雨燕類(Strisores)、 鴿鴇類(Columbaves)、鶴形目(Gruiformes)、水濱鳥 類(Aequorlitornithes)和望外鳥類(Inopinaces;包含鷹形目 Accipitriformes、鴞形目 Strigiformes等)五大類群(圖二)。較晚演化出來的望外鳥類大多屬於半晚熟鳥或晚熟鳥,其中,麝雉(Opisthocomus hoazin)則是唯一的例外, 屬於半早熟鳥,但同時也是最原始的望外鳥類。因此從演化樹可得知,在望外鳥類出現時發生了一次從早熟鳥到晚熟鳥的演化事件。
在新鳥類中,夜鷹及雨燕類和鴿鴇類是兩個較古老的陸棲鳥類類群。夜鷹及雨燕類雖然是早期就演化出來的類群,但所有成員都屬於半晚熟鳥或晚熟鳥,這表示最早的晚熟模式演化事件就已發生在這個早期就演化出來的類群中。鴿鴇類則涵蓋所有四種發育模式,其中有鴿型類和鴇型類兩個不同的演化枝,各自衍生出了晚熟鳥種類,顯示晚熟模式演化事件在這個類群中至少獨立發生了兩次。 其餘的兩大類,鶴形目和水濱鳥類,大多是水禽或是飛行能力不佳的陸禽。所有的鶴形目都是水禽且屬於早熟鳥類,根據演化樹可知(圖二),鶴形目可能是水濱鳥類的祖先。水濱鳥類的發育模式則呈現高度多樣性,其發育模式演化是遵照早熟鳥至晚熟鳥的順序,類似於整體鳥類的演化趨勢,可以推估此演化枝系中至少發生了一次晚熟鳥演化事件。
表一、雛鳥孵化模式的辨識特徵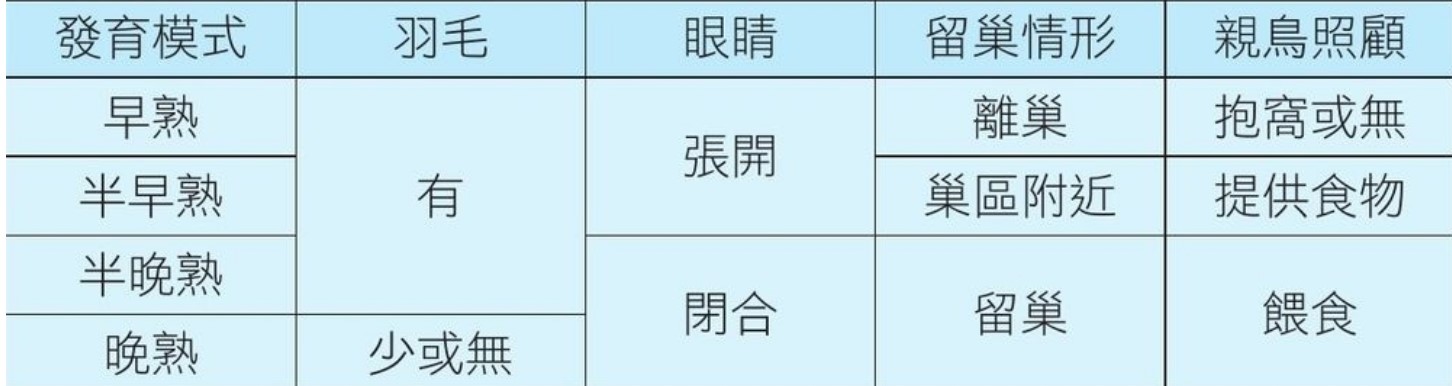
此表是由(Chen et al., Zoological Studies 58:24, 2019)修改而得。
總結而言,圖二顯示在現生鳥類演化過程中,至少有五個獨立的晚熟模式演化事件。
鳥類演化過程中新生絨羽的演化趨勢
新生絨羽是辨識「早熟—晚熟頻譜」的特徵之一,絨羽可維持雛鳥的體溫,是鳥類在演化上為了適應多變的環境而得。晚熟鳥和早熟鳥之間新生絨羽的發育差異,可能是羽毛的巨觀和微觀發育樣式的差異所致。
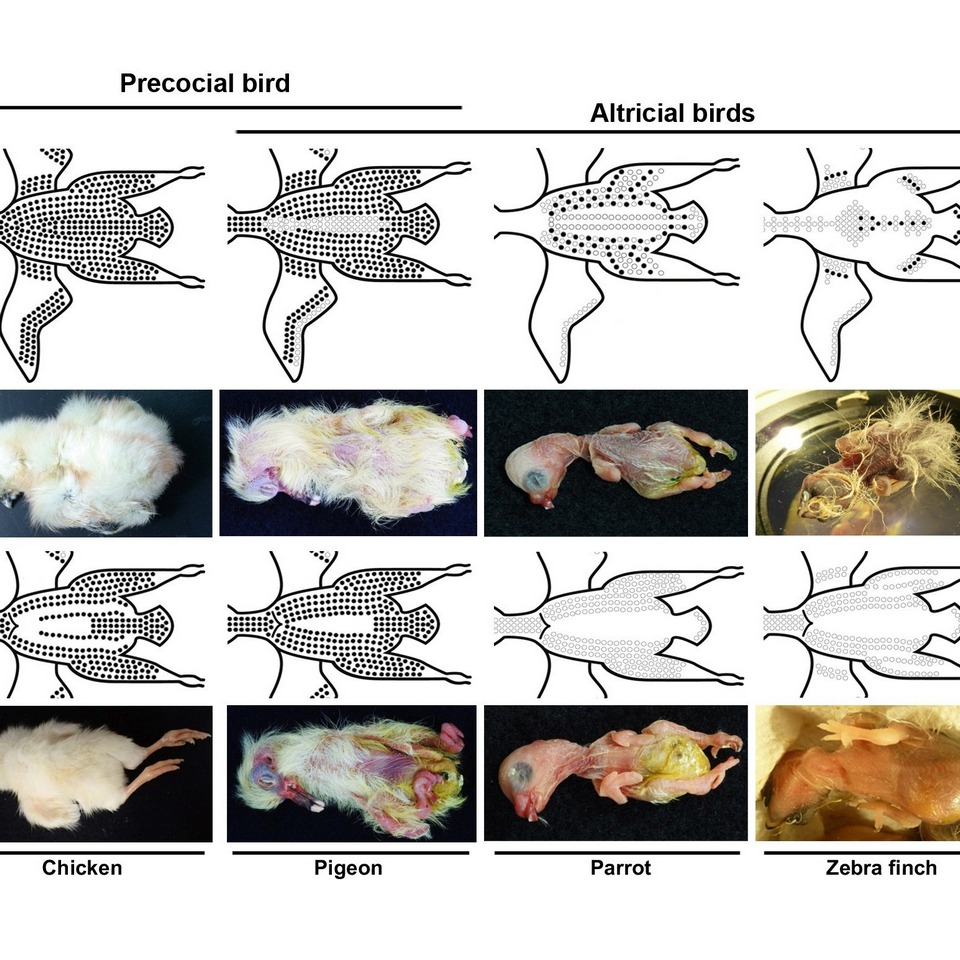
羽毛巨觀發育樣式是指雛鳥「有羽區」與「無羽區」的分佈。羽毛的毛囊集中生長在「有羽區」,周圍則被裸露的「無羽區」隔開(圖三)。晚熟鳥的有羽區面積比早熟鳥的小,舉例而言,由於雞是早熟鳥,雛雞的新生絨羽幾乎覆蓋了整個身體表面;斑胸草雀(zebra finch)則是晚熟鳥,孵化時有羽區僅覆蓋了身體的一小部分(圖三)。無羽區的演化有三種可能解釋:(1)無羽區可減輕羽毛的總重量、(2)裸露區域(無羽區)可使鳥類更適應身體和羽毛的運動、(3)裸露區域可幫助晚熟鳥在飛行或育雛過程中調節體溫。晚熟鳥通常有比較好的飛行能力,換句話說,羽毛的巨觀發育樣式與飛行能力演化有關。
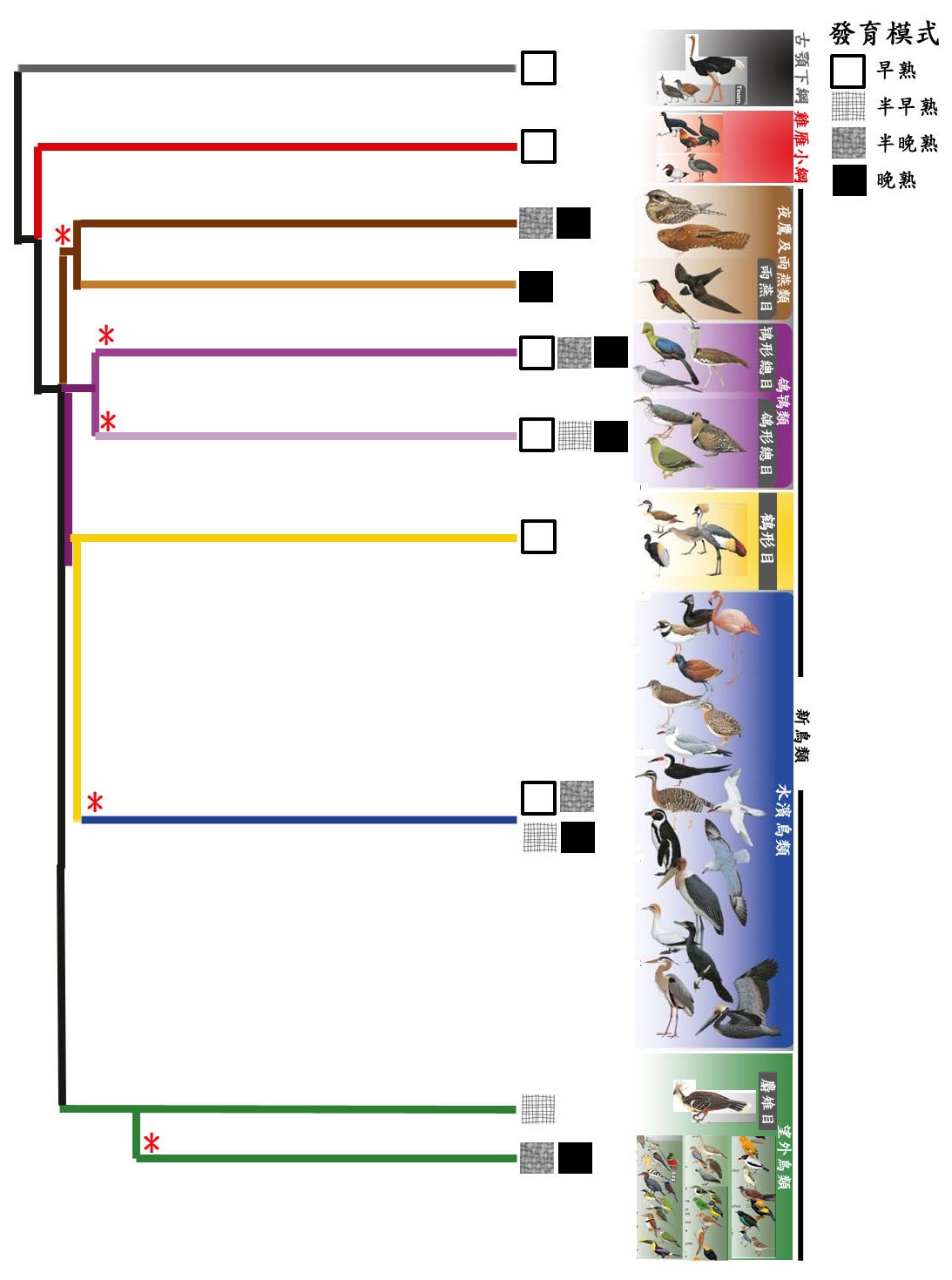
圖二、鳥類演化樹以及相對應的四種主要雛鳥發育模式。此圖是由
(Prum et al., Nature 526: 569-573, 2015)修改而得。「*」代表是發生過一次從早熟鳥演化到半晚熟鳥或晚熟鳥的事件
羽毛的微觀發育是指單個羽芽的發育類型。羽毛的多樣性,可以在不同發育階段,或是在成鳥的不同身體區域觀察到。飛羽可利於飛行,廓羽可以護身與塑身,而絨羽可以保暖。在鳥類發育上,大多數鳥類孵化時只有新生絨羽,隨著成長的階段,正羽逐漸取代新生絨羽。我們發現雞和斑胸草雀的新生絨羽具有不同的生長模式(圖三), 這顯示鳥類的演化可能和羽毛微觀發育模式的變化相關。
為了瞭解各種新生絨羽如何影響鳥類演化,我們回顧了相關的文獻,聚焦在主要四種(早熟、半早熟、半晚熟、晚熟)發育模式的異同。
鴯鶓(emu)是古顎下綱的一員,屬於早熟鳥(圖二)。與雞相比,鴯鶓的卵更大,需要較長的時間孵化。此外,雛鳥體型更大,並發展出獨特的前肢。但是,與雞相同,鴯鶓的幼雛的全身都覆蓋著新生絨羽,孵化後不久即可自行覓食。憑藉這些觀察顯示,早熟鳥具備相似的新生絨羽分佈和進食行為。
鴿子(pigeon)是屬於鴿鴇類,其發育模式仍有爭議。一些研究將其歸類為晚熟鳥,另一些研究則將其歸類為半晚熟鳥。儘管鴿子的幼雛需要親鳥較多的照料,但是絨羽覆蓋了幼雛大部分的身體表面(除了後腹側區域之外)(圖四)。因此,我們將鴿子歸類為半晚熟鳥。
斑胸草雀(zebra finch)屬於鳥類譜系最多樣的雀形目(Passerineform)。所有的雀形目皆是晚熟鳥。斑胸草 雀的幼雛的有羽區有兩種新生絨羽的發育模式:第I型羽芽直接從毛囊生長出正羽,而沒有新生絨羽的階段,導致在孵化時該區域皮膚裸露。第II型羽芽則像早熟鳥的羽芽一樣,首先發育出新生絨羽,而後被同一毛囊中生長的正羽取代。兩種類型羽芽的正羽皆在相近的時間點發育(D7, 圖三)。
虎皮鸚鵡(budgerigar)屬於雀形目的姊妹支,也是 一種晚熟鳥,但是牠們新生絨羽的發育與斑胸草雀不同, 這兩個物種的幼雛均沒有腹部絨羽。至於體背上,斑胸草雀幼雛已有成熟的絨羽,但鸚鵡幼雛的絨羽卻仍在發育中(圖四)。我們只能在鸚鵡幼雛腹部觀察到跟斑胸草雀一樣的第I型絨羽發育,其背部的絨羽發育機制可能和斑胸草雀的不同。
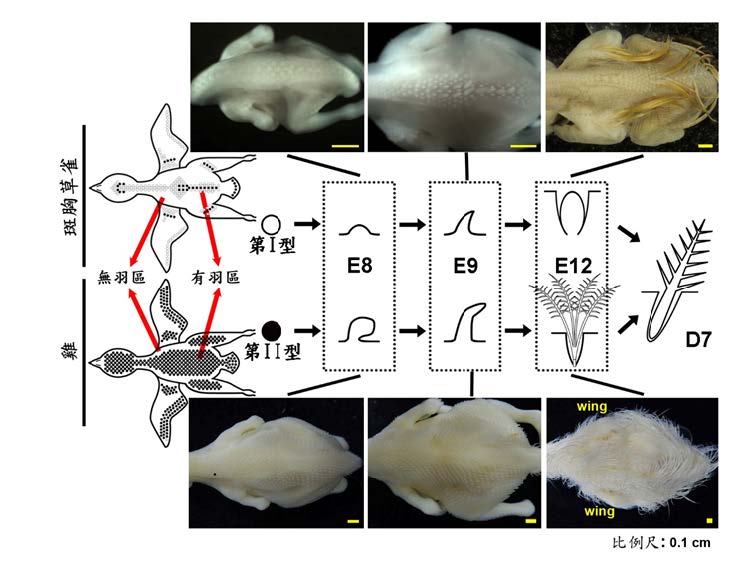
圖三、斑胸草雀跟雞的「有羽區」及新生絨羽發育差異的示意圖。斑胸草雀胚胎羽芽顯示了兩種絨羽發育模式:第I型發育模式不會長出絨羽(空白部位),而第II型發育模式(黑色圓圈部位)會長出絨羽。兩種發育模式的羽芽最後都會長出正羽。雞胚胎所有的羽芽都是第II型發育模式。E8、E9、 E12分別是指胚胎發育的第8、9、12天。D7是指孵化後第7 天。各圖中的比例尺是指 0.1公分。 此圖是由(Chen et al., Zoological Studies 58:24, 2017)修改而得。
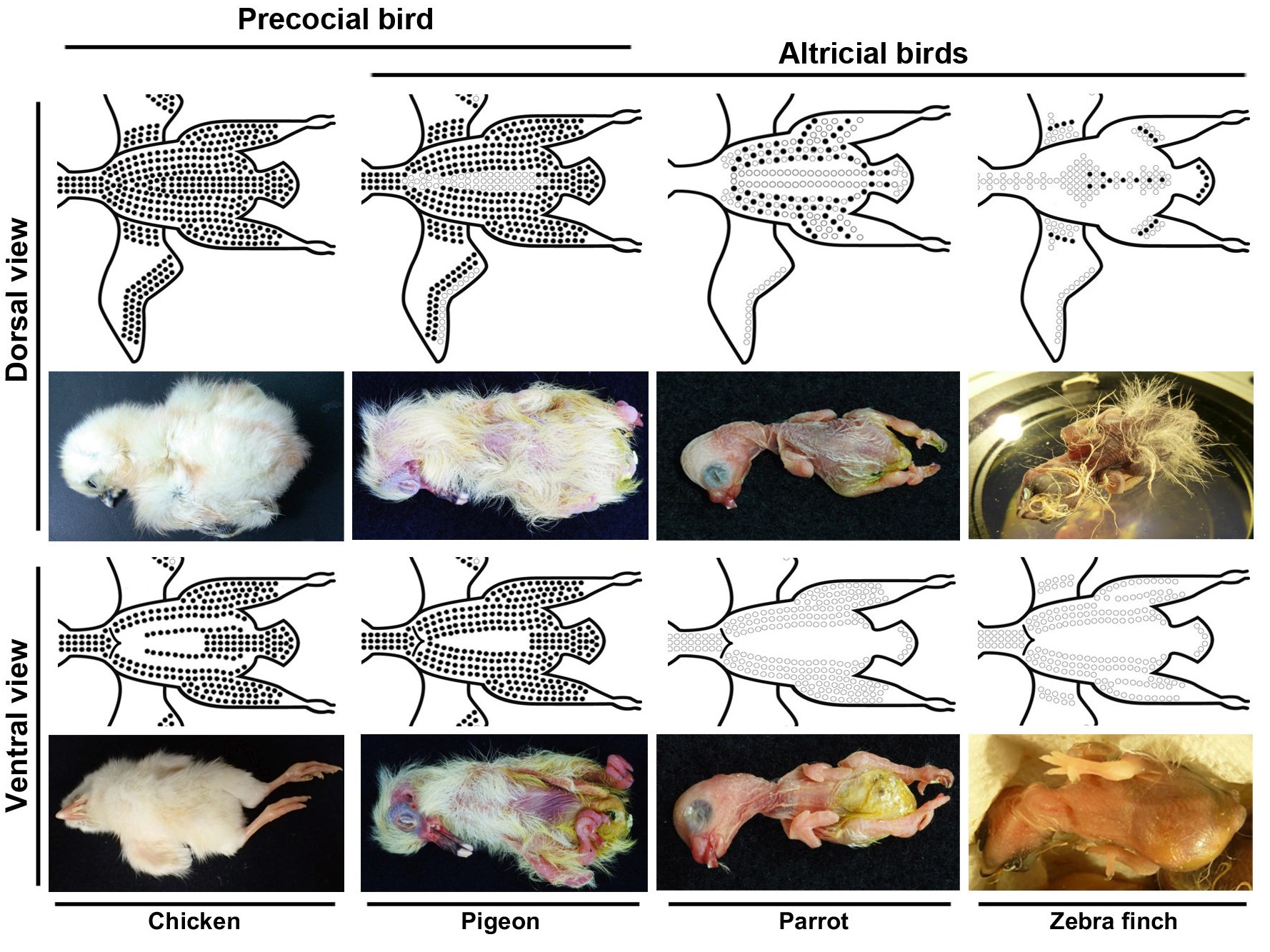
圖四、鳥類幼雛及其示意圖。雞(早熟鳥)、鴿子(半早熟鳥)、鸚鵡和斑胸草雀(晚熟鳥)的背視角(dorsal view)(上排)以及腹視角(ventral view)(下排)。黑色圓圈部位會長出絨羽。此圖是由(Chen et al.,
Zoological Studies 58:24, 2019)修改而得。
通過比較上述四種鳥類的新生絨羽發育,我們提出在鳥類演化中,新生絨羽的演化過程:新生絨羽最初覆蓋整個早熟鳥幼體。在早熟鳥類占領了大部分陸地生態棲位後,具備更佳飛行能力的半早熟、半晚熟鳥才開始出現並將其棲息地擴展到水域、海洋或高於地面的地方。半早熟及半晚熟鳥可在隱蔽的地方築巢,因此牠們的幼雛可以待在巢中生長發育,而不再需要腹側的絨羽保持腹部體溫。最後,晚熟鳥開始出現,牠們具有較高的智力並可以建立複雜的巢,提供更佳的保護,因此可以縮短雛鳥的孵化時間,並將能量重新分配給其他器官(例如消化器官)藉以加速孵化後的發育。
結論與展望
「早熟—晚熟頻譜」描述鳥類出生時的發育模式,而發育模式與親鳥的照料、棲息地的選擇和環境適應高度相關。新生絨羽是「早熟—晚熟頻譜」的辨識指標之一,並且與鳥類演化有密切的關係,但過去的研究欠缺大尺度的分析。
由於鳥類在白堊紀大滅絕(K-Pg extinction)後快速地輻射分化,其演化樹一直難以精確構建。完整的基因體序列有助於建構精確的演化樹,因此為了回答這個問題, 科學家根據48種鳥類的全基因體構建了鳥類的演化樹。然而,物種演化樹的構建不僅取決於基因體的測序完整度,還取決於代表性物種的種類。我們所使用的鳥類演化樹(圖二)囊括了所有主要鳥類譜系,但是基因體測序完整度較低。雖然與由較完整的基因體序列但包含較少的鳥類譜系構建而成的演化樹相較之下,顯示出些許差異(例如,猛禽的親緣位置),但是兩者都適用於我們建構的絨羽演化模型。當然鳥類的演化樹可能很快就又會被修改, 例如最近在雀形目中進行的大規模基因體學研究揭示了更精確的演化樹和演化軌跡。因此若能對基因體序列相對不完整的半早熟和半晚熟物種進行更完整的測序,將能增進「早熟—晚熟頻譜」的演化樹正確性。此外,積累形態描述的紀錄,也可能會改變我們的模型,由於大多數形態學數據來自野外鳥類的調查和記錄,所以很難判斷現有的數據是在鳥類幼體孵化後哪一個時期所記錄的。而我們已經觀察到,至少在鸚鵡中,出生後新生絨羽會急劇增長,因此定義頻譜應該針對剛孵化的狀態進行判定。
羽毛發育涉及許多分子的相互作用,目前不同發育階段的主要調控因子基本上已經被研究清楚,但是其上游的調節因子和機制仍然尚待釐清。有關此議題的討論,可以參見Chen et al.(Zoological Studies 58:24, 2019)。
羽毛發育和演化的研究,開拓了我們對於複雜結構如何增進生物續存的知識。但是無論在型態或是身體上的分布,新生絨羽多樣性的重要性常常被低估。為了適應環境,有些鳥類從早熟鳥演化到晚熟鳥,然而大多數的有關早熟鳥與晚熟鳥的鑑別特徵難以定性或量化,因此長久以來難以精確地定義它們在「早熟—晚熟頻譜」的位階。新生絨羽則是辨識特徵中唯一可以定性及定量的,因此,我們對「早熟—晚熟頻譜」的演化重新做了推論。
此外,不同發育階段的鳥類個體表現出不同類型的羽毛。當幼鳥準備離巢時,大部分的新生絨羽會被替換成為正羽和飛羽,使牠們得以飛翔。而經過幾次換羽後,毛囊會衍生出具有更多功能的羽毛,以便為成鳥所用,例如偽裝、遷徙、越冬與求偶等。大多數鳥類僅會發生一次從新生絨羽替換到正羽的過程,但是替換的機制從未被仔細研究過。因此,羽毛替換的研究將有助於我們探索鳥類「早熟—晚熟頻譜」演化的奧秘。
後 記
絨毛在新生雛鳥身體上的分布是區別早熟鳥和晚熟鳥最好的指標。近年來分子生物學及基因體學的發展使我們對絨毛的演化有了相當的瞭解,在鳥類的演化史上從早熟鳥到晚熟鳥的演化至少發生過五次。但目前對絨羽發育的分子機制瞭解有限,是有待解決的問題。
作者簡介

李文雄
中央研究院生物多樣性研究中心特聘研究員,Brown大學應用數學博士,專長為演化、基因體學和生物資訊。研究範圍涵蓋分子演化、鳥類演化、基因體演化及人類演化。2003年被選為美國國家科學院士,並獲得 Balzan Prize for Genetics and Evolution。喜好音樂、文學、旅行和世界新聞。經常去體育館。

陳志冠
美國南加州大學凱克醫學院博士後研究員,臺灣大學生演所博士。專長為演化發生學及生物資訊,以及養鳥等小動物。研究領域包含演化生物學、發育生物學、次世代定序技術、表觀基因體學等。目前主攻羽毛的演化與發育,期待能解開更多羽毛之謎。研究之餘喜歡運動、旅行、冷知識及飼養小生物。

常見的一些早熟鳥、半早熟鳥、半晚熟鳥和晚熟鳥。